
The impact of helminth infections associated with the lack of efficacy of the major commercial compounds and reports of anthelmintic resistant strains worldwide [1,2], has raised the need to find alternative methods for the control of parasites of equids. Moreover, the low probability of obtaining new synthetic anthelmintic drugs [3], and the increasing demand for organically produced food [4], represent additional factors that further increase the need for sustainable methods for parasite control.
More recently, new approaches have been widely studied for the control of equid nematode infections. These include pasture management [5,6], the selection of adapted/rustic and resistant breeds [7], biological control methods [8,9], and the offering of nutraceuticals and other natural therapies [10,11,12]. The latter are considered particularly promising for the control of gastrointestinal nematodes (GIN) in livestock [10,13]. In fact, natural compounds can be the source of new anthelmintic molecules or can be directly used for the formulation of food supplements [14].
People in most regions, have always used plants and other natural compounds as treatment for various ailments and diseases, including those caused by parasites [15,16]. Among the countless natural compounds known for their benefits, marine compounds have also shown a wide range of activities such as anthelmintic, antibacterial, anticoagulant, antifungal, antiprotozoal, and antiviral [17,18]. Due to their nutritional value and the content of bioactive molecules, especially marine macroalgae (seaweeds) are often used as food, herbal medicines, and dietary supplements [19]. Seaweeds, comprising thousands of species, are vegetal organisms empirically distinguished in red, brown and green algae belonging to three phyla: Rhodophyta, Ochrophyta and Chlorophyta, respectively. Such phyla, however, differs each other in addition to pigmentation, more substantially in thallus structure, biochemical characteristics including photosynthetic pigments, reproductive structures, ultrastructure of chloroplasts, storage compounds, etcetera. Red algae are known not only for their use in food and cosmetic productions [20], but also for their numerous biological activities, including anticancer [21], antidiarrheal [22], antimicrobial and anthelmintic activities [17,22,23,24]. The latter activity has been known since ancient times in Eastern countries and around the Mediterranean area including Italy, where red algae were used as an anthelmintic in popular medicine [25,26].
The anthelmintic activity of red algae was previously reported in Italy, such as Ellisolandia elongata (syn. Corallina elongata), taken as decoction in the Tuscany Archipelago [27], and Hypnea musciformis eaten fresh for the same purpose used in the coastal of Abruzzo [25,26]. In Sicily, many species of red algae are also well known for their medicinal property [28]. During an ethnobotanical survey carried out in the Sicilian Egadi Islands, several reports of the anthelmintic use of an aqueous extract (about at \(10%\)) made with a mixture of red algae gathered on the rocks in intertidal zone, which was called “semenza de vermi”, were collected. This aqueous extract was widely used in the past mainly to treat worm infections in children, and it was even marketed out of the islands until the post-war period (Laura Cornara, unpublished data).
Considering the ethnobotanical relevance of this remedy used for the treatment of human worm infections in the Italian folk medicine, this study was aimed to assess in vitro the effective anthelminthic properties of this red algal aqueous extract on donkey nematodes. Another objective was to identify the components of the red algal species that composed the mixture of this remedy. In fact, the confirmation of the anthelminthic properties of this extract could be extremely useful for its possible use as a new tool for the control of nematode infections in donkeys, and possibly be used in other mammals.
We collected a mixture of red algae on sea rocks in the intertidal zone, following the method used by elderly people from Egadi Islands to prepare anthelmintic remedies. The material was collected at Favignana island, Egadi islands, Sicily, Italy, in January 2017. The algal species in the mixture were identified according to Furnari et al., [29], and Serio et al., [30] description keys\(’\) (see Figure 1).

From this mixture, a \(10%\) aqueous extract (red algae extract, RAE) was obtained. For this, \(50\) g of dried algae were macerated in \(500\) ml of distilled water for \(3\) h, followed by sonication for \(30\) min. The crude extract was filtered (\(0.45 \mu m\) filter), frozen and lyophilised (yield of \(75.6%\) on a dry matter basis). The dried extract (\(10\) g) was solubilized in distilled water (\(100\) ml). The obtained solution was frozen until it was used for the in vitro activity tests.
In the red algae mixture, Palisada tenerrima, Laurencia intricata and Laurencia spp. were identified (see Figure 1). Moreover, in the collected materials, P. tenerrima, L. intricata and Laurencia spp. were found in the proportion \(2:1.5:0.5\), respectively.
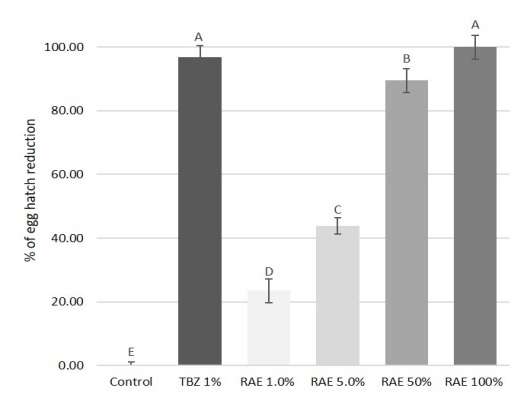
The pure RAE was able to cause \(100%\) inhibition of egg hatch, showing an efficacy comparable to TBZ \(1% (98.7%)\) (see Figure 2). Furthermore, the anthelmintic efficacy of the extract was found to have a concentration-dependent effect, with an equation of \(y = 26.35x – 27.99\) and the correlation coefficient \((R2)\) of \(0.968\). In fact, at all other dilutions (\(50, 5\) and \(1%\)), the RAE tested in the study showed a decreasing efficacy of \(89.5, 43.7\), and \(23.4%\), respectively. Moreover, the two lowest dilutions of the extract showed a significant \((P < 0.05)\), lower activity than the pure extract and the reference drug.
The identification of GIN genera from the pooled faecal cultures revealed a high prevalence \((>90%)\) of cyathostomins. More specifically, we found species belonging to the genera Cylicocyclus and Cylicostephanus. The remaining \(10%\) of the identified larvae were identified as Triodontophorus.
We have demonstrated the effect of RAE against GIN eggs of donkeys. Red algae are a group of eukaryotic organisms also known as rhodophytes, most of which live in warm seas [36]. About \(4000\) different species of red algae have been identified [37]. The alimentary use of red and other algae is particularly widespread in Eastern countries, where they are considered a gastronomic delicacy and a food source with excellent nutritional qualities (i.e. proteins, vitamins) [38,39]. Algae are also used as stabilizers, thickeners, emulsifiers, and gelling agents, and as adjuvants in the treatment of obesity, cellulite, and related diseases [18,20]. However, nowadays green, brown, and red marine algae are also considered an important source of bioactive compounds that can be used to inhibit pathogens, such as bacteria, viruses, and parasites [17,40,41,42,43], including helminth infections [17,41].
Digenea spp. red algae are in fact known for more than a thousand years for their anthelmintic properties against intestinal ascarid worms [44,45]. Kainic acid is considered the active compound responsible for the deworm activity of this group of algae and it is probably due to its ability to induce a neuromuscular paralysis, which interrupts the peristaltic movements of the intestine of the parasites [23]. Kainic acid belongs to the family of kainoids that include also domoic acid, another neurotoxic compound produced mainly by marine planktonic diatom algae [46]. The similar activity of domoic and kainic acids is due to their related structure with glutamic acid, which alters the function of some ion channels [45,47]. At the dose of \(5\) to \(10\) mg, the domoic and kainic acids can permit the expulsion of roundworms without causing any side-effects in the infected hosts [23,48]. Domoic acid may be concentrated in marine seafood, such as oysters, mussels and crustaceans. Sea-food poisoning can be related to domoic acid to fish and humans [49,50]. The ingestion of marine organisms containing high levels of domoic acid, may even cause the death of fish and other marine mammals.
A further major class of metabolites produced by marine algae are represented by terpenes [43]. The Rhodophyceae are characterized by a high content in polyhalogenated monoterpenes showing potent and wide antimicrobial activities [51,52]. In this study, the \(10%\) aqueous extract from P. tenerrima, L. intricata and Laurencia spp. red algae showed a high anthelmintic efficacy against GIN of donkeys, comparable to that of the reference drug (TBZ \(1%\)). In addition, the in vitro anthelmintic efficacy showed a concentration-dependent effect against GIN eggs. Even the efficacy of RAE at \(50%\) was high, with an egg hatch reduction of about \(90%\).
This is the first study in which the anthelmintic activity of a RAE was tested using the EHT against nematode eggs of GIN of donkeys. It is interesting to note that the anthelmintic properties of algal species of the genus Laurencia were already reported [41,53]. Indeed, among \(11\) sequiterpenes and a long chain aldehyde from Laurencia dendroidea (syn. Laurencia scoparia), a halogenated \(\beta\)-bisabolene sesquiterpenoid showed anthelmintic properties against Nippostrongylus brasiliensis [41], a gastrointestinal nematode of rats with a life cycle and morphology similar to the human hookworms [54]. Moreover, very similar results to those obtained in the present study were reported when evaluating the anthelmintic activity against Meloidogyne javanica, an important plant root-knot nematode species. The methanolic and aqueous extracts of several seaweed species were tested using the EHT and the larval mortality test [55]. More specifically, in the previous study, a \(10%\) aqueous extract of the algal species Sargassum tenerrimum, Padina tetrastromatica and Melanothamnus afaqhusainii were able to cause a reduction of about \(96%\) of the egg hatch of M. javanica [55]. However, in the study of Kha et al., [55], the \(10%\) aqueous extract from the algal species, including Laurencia pinnatifida (now Osmundea pinnatifida), showed a lower anthelmintic activity in the EHT when compared to the \(10%\) aqueous extract examined in the present study.
Nevertheless, the phytochemical profile of the RAE tested in this study was not performed, and further studies aimed to identify the different components present in the extract are needed. In fact, the knowledge of the main bioactive components of this aqueous red algal extract could allow to estimate its potential toxic effects to mammals and, therefore, its effective potential as an anthelmintic of seaweed origin. Nevertheless, it is plausible to hypothesize that the anthelmintic activity of the RAE can probably be ascribed to the presence of kainic acid, as reported in other species of red algae [44,45]. Other compounds may also be important (i.e. sesquiterpenoids, \(1\)-aminocyclopropane-\(1\)-carboxylic, and acrylic acids) to red algae, which in previous studies were found to possess anthelmintic properties [53,55].
Further studies are needed to evaluate the anthelmintic activity of each of the three algal species contained in the mix of algae tested in this study. In fact, these latter studies may permit to assess whether there is a synergism among these species, or if one species has higher activity than the others. Finally, although in previous studies nematodes have been found to be highly tolerant to salinity [56,57], the potential anthelmintic effects of the content in sodium chloride of the tested algal extract cannot be excluded.
In conclusion, results from this study seem to confirm the anthelmintic properties previously reported in the Italian folk medicine for the red algae extract. Further studies based on the bio-guided purification of the red algae extract are encouraged.